Холестерол синтезинин жөнгө салуу
холестерол синтези жөнгө салуу - анын негизги энзим (ГМГ КоА) ар кандай жолдор менен созулган.
Phosphorylation / dephosphorylation ГМГ-редуктаздык. инсулин үлүшүн бир топ көбөйтүү менен / Бул энзим phosphorylated жана активдүү болуп жатат глюкагон. 2 энзим аркылуу инсулин иш-аракет.
Креатинкиназа ГМГ-КоА-phosphatase, алардын privraschaet Креатинкиназа жигердүү dephosphorylated мамлекет:
Phosphatase ГМГ-КоА-dephosphorylated жигердүү мамлекет аны айландыруу менен редуктаздык. Эгер ушундай болуп жатса натыйжасы активдүү түрү defosorilirovannoy ГМГ-КоА-түзүү болуп саналат.
Ошондуктан, холестерол absorptive мөөнөтү өскөн. Бул мезгил ичинде өсүш жана холестериндин синтезине булагы субстрат бар учурунда - acetyl - КоА (acetyl-КоА-глюкоза жана май кислоталарынын чиришинен пайда болгон эле нан бар карбонгидрат жана майлардын, натыйжада).
аркылуу глюкагон пост absorptive абалда proteingenazu А стимулдайт phosphorylation боюнча ГМГ - КоА - редуктаздык, жигердүү эмес мамлекет аны айландыруу. Бул иш-аракеттер да глюкагон бул phosphorylated жигердүү абалда ГМГ-КоА-сактоо, phosphorylation жана phosphatase ГМГ-КоА-жылдын inactivation сигнал улам жардам берет. Постту absorptive орозо мезгилинде холестерол айкалышынын натыйжасы катары жана ички синтези басат. тамак-холестерол мазмуну 2% га чейин эске алынган болсо, анда чукул ички холестерол синтезделишинде азайган. Бирок, холестерол синтездөөгө эч кандай толук токтотуу жок.
ар кандай адамдардын тамак холестерол аракетине холестериндин биосинтези тыюу салуу даражасы ар түрдүү болуп саналат. Бул холестерол түзүү жараяндардын инсандардын көрсөтөт. холестерол синтези абалын көрyп азайтуу, ал канга анын топтоо кыскартуу мүмкүн.
Эгерде бузулган холестерол тамак- аштын ортосундагы тең салмактуулук жана анын метаболиттери денеде бир тараптан жана өт кислоталары чыгарууда жана холестериндин - башка, Кан менен кыртыштарда холестерол топтолуу өзгөрөт. абдан оор кесепеттерге алып келсе, холестерол жана кан топтолуу жогорулашы менен байланышкан (hypercholesterolemia) бул атеросклероздун жана cholelithiasis ыктымалдыгын жогорулатат.
Үй hypercholesterolemia (НСН) - бул түрү көбүрөөк кездешет - 200 кишиге 1 иши жөнүндө. hypercholesterolemia менен тукум куума кемтиги клеткалар тарабынан LDL өздөштүрүү бузуу менен, демек, LDL жана катаболизм кыскартуу болуп саналат. Натыйжада, кан LDL бир LDL кандагы холестеролду, анткени анын өкчөмө таш боюнча топтолушун жогорулатат-жылы. Ошондуктан, артериялардын капталдарында жылы GCN кыртыштарда холестерол мүнөздүү катмарланышы, айрыкча, тери менен (xanthomas) болот.
ГМГ-КоА-ажыратуу
холестерол зат алмашуу жолун акыркы продукт. ген ГМГ тако ыкма өз синтезин жокко КоА-редуктазы буйруйт ылдамдыгын азайтат. боор жигердүү холестерол, өт кислоталар жана ошондуктан үгүттөө ген ГМГ-КоА ишинен өт кислоталарынын синтези жүрөт. ГМГ-КоА-3 жөнүндө синтези кийин бар, бул энзим холестерол синтези ажыратуу натыйжалуу жөнгө салуу болуп саналат.
Сиз издегенибизди табууга эмес беле? издеп көргүлө:
Холестерол Esters алмаштыруу
холестерол Foundation эркин холестерол жана холестерин Esters камтыйт, Бегаим буудай да клеткаларда жана кан липопротеин бар.
II бөлүм. Зат алмашуу жана энергия
холестерол Колдери клеткалары acyl-КоА-ho- lesterin acyltransferase (АСАТ,) менен иш-аракет менен пайда болот:
Acyl-КоА-+ холестерол - * HS-дулгуяк + Atsilholesterin
Адам клеткаларда, олуттуу linoleilholesterin түзүлгөн. өтө аз санда клетка кабыкчасындагы айырмаланып эркин холестерол жана анын ТББнын жана негизинен көрбөйбүз тамчыларын бөлүгү катары Органеллдер бар. Клеткадагы холестеролду сактоо үчүн механизм катары - Эстер түзүү механизми, экинчи жагынан, ал эми кабыкчасы ашыкча холестерин жок эле, бир жагынан, көрүүгө болот. камдардын тартуу esterase энзим деп hydrolyze холестерол ТББнын катышуусу менен пайда болот:
Atsilholesterin + H 2O - * F кислотасы + холестеролду irnaya
Синтези жана Esters гидролизи көп клеткалардан, бирок, өзгөчө ар түрдүү, бирок adrenocortical клеткаларында пайда болушу: Бул клеткалар ар холестерол Esters 80% га чейин, ал эми башка клеткаларда адатта 20% дан кем эмес, алдына койду.
Жылы liponroteinah кан Эстер түзүү lecithin-холестерол acyltransferase (LCAT) катышуусу менен кездешет холестериндин үчүн lecithin боюнча acyl калган (мен-абалын өткөрүп катализатор (сүрөт. 10.31). LCAT боордо өндүрүлгөн, канга бөлүп, липопротеиддердин бекитилип берилет. ар кандай липопротеиддердин үчүн Колдери баасы кыйла айырмаланып турат жана Mardzhan-Splendens полипротеиндерине көз каранды, LCAT жандандыруу (негизинен Тайна-жана CI) же тоскоол болуучу (C-II) бул энзимди. Тайна-AT үлүшү болгон липопротеинди абдан активдүү LCAT каттоо olee бүт белоктордун 2/3. oleic жана линол кислоталарынын Esters пайда болгон ири таякчасынын наамдарга татыктуу болгон. Эстер калыптандыруу башка липопротеиддердин липопротеинди караганда жайланган калып менен пайда болот.
Сүрөт. 10.31. LCAT аракетине холестерол Esters түзүү
LCAT phospholipid monolayer жайгашкан суб катмар холестерол сыяктуу липопротеинди жана пайдалануунун жер катмарынын, локалдашкан. толук бакырык улам бул жерде пайда болгон холестерол ТББнын начар сакталып
10-бөлүм: бөлүшүү жана липиддерди камтыган иш-милдети
phospholipid monolayer жана липопротеин менен көрбөйбүз ядродогу жатышат. phospholipid monolayer клетка кабыкчасындагы же башка lipop roteinov тартып холестерин менен толтура алышат холестерол үчүн орун бошоткон адаты. Ошентип, LCAT HDL иш-аракеттердин натыйжасында lovush Кой холестерол бар.
Өт кислоталарынын синтези
холестерол боор өт кислоталары динин кабыл алат. F сен cholanic кислотасы туунду (сүрөт. 10.32) катары каралышы мүмкүн муздап elchnye.
сыяктуу Cholanic кислотасы денесинде пайда болгон эмес. холестерол тартып hepatocytes менен түздөн-түз chenodeoxycholic алган, сени муздап cholic - негизги Өт кислотасы (Figure 10.33, ошондой эле сүрөттү кара 10.12 ...).
Холестериндин биосинтези
Холестериндин биосинтези эндоплазма торчосу болот. молекуласында баары көмүртек атомдору булагы, ошондой эле май кислотасынын синтези менен acetyl SKoA курамы бул митохондриялык Киириитэ кирген болуп эсептелет. холестериндин биосинтези 18 ATP жана NADPH 13 молекулаларды арнаган.
Cholesterol билим берүү бир нече этаптан топтоштурууга болот 30дан ашуун сезимге болуп саналат.
1. mevalonic кислотасынын синтези.
биринчи гликогенолиз, зээр эки синтезте, ал эми 3-Гидрокси 3-methylglutaryl-SKoA натыйжалуу энзим болуп синтези кийин hydroxymethyl-glutaryl-SKoA редуктаздык (SKoA ГМГ-редуктаздык) mevalonic кислота түзүү.
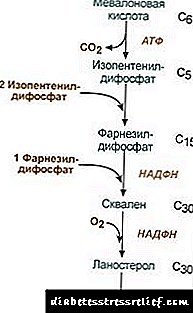 |
Схема холестеролунун синтезте2. Synthesis izopentenildifosfata. mevalonic кислотасы тиркелет, бул этап үч кездешүүчү калган учурда, анда ал decarboxylated жана денеси суусузданып кеткен. 3. izopentenildifosfata farnezildifosfat синтезделген үч молекулалар биригишинен кийин. 4. squalene синтези эки farnezildifosfata милдеттүү калдыктарынан пайда болот. 5. комплекстүү мамиле кийин lanosterol үчүн squalene cyclizes сызыктуу. 6. кошумча метил топторду, калыбына келтирүү жана молекула холестерол үчүн сөздүн isomerization алып салуу. Hydroxymethylglutaryl-S-КоА ишин жөнгө салуу3. холестериндин биосинтези курсу да топтоо боюнча көз каранды белгилүү бир берүү белокБерүү милдеттүү жана метаболиттерди синтези hydrophobic арачылардын транспорттук. Сиз сурап же пикирин калтыра алат. жөнгө салуунун негизги пункту mevalonic кислотасы пайда жооп болуп саналат. 1. аллостерикалык жөнгө салуу. Холестерол жана боор - Өт кислотасы жана үгүттөө ГМГ-КоА. 2. ГМГ-КоА-жылдын Репрессиялар синтези холестеролду редуктаздык. 3. ГМГ-КоА-жылдын phosphorylation-dephosphorylation менен жөнгө салуу активдүү unphosphorylated түрү болуп саналат. кубулуштардын татаал каскадын менен жандануусуна - глюкагон өчүрүү жана инсулин себеп болот. Ошентип, холестерол синтезинин курсу пост absorptive жана absorptive мамлекеттерди өзгөртүү менен өзгөрөт. 4. боордо ГМГ-КоА-айкалышынын баасы так күндүзү ёзгёръълёргё дуушар болгон: Түн ортосунда максималдуу жана минималдуу - эрте. холестерол Esters алмаштыруу клеткалар иш-аракет менен холестерин Колдери жүрөт acyl-КоА-холестерол acyltransferase (АСАТ): Acyl-КоА-+ холестерол ® HS-КоА-+ Atsilholesterin клеткалар негизинен linoleilholesterin түзүлгөн. Esters негизинен көрбөйбүз тамчыларын курамына Органеллдер кездешет. Эстер пайда көргөн, бир жагынан, кабыкчасы ашыкча холестериндин жоюу үчүн бир механизми катары, экинчи жагынан, ал эми мүмкүн - Клеткадагы холестеролду сактоо үчүн бир механизм болуп калды. камдардын тартуу энзимдердин катышуусу менен болот esteraseхолестерол Esters гидролиз: Atsilholesterin + H2On ® Cholesterol + май кислотасы ТББнын синтези жана гидролизи үстү кортексинин клеткалары өзгөчө активдүү болот. кан липопротеиддердин Эстер пайда катышуусу менен болот lecithin-холестерол acyltransferase (LCAT) холестерин менен lecithin бир acyl өмүрдөн берүүгө көмөктөшөт. LCAT боордо өндүрүлөт, канга бөлүп, липопротеиддердин тиркелет. ал жер катмарынын локалдашкан LCAT липопротеинди, абдан активдүү. холестерол hydrophobic ТББнын жерде пайда болгон жана көрбөйбүз ядродогу чөмүлдүрүү турат. phospholipid monolayer клетка кабыкчасындагы же башка липопротеиддердин тартып холестерин менен толтура алышат холестерол үчүн орун бошотту. Ошентип, LCAT HDL иш-аракеттердин натыйжасында холестерол тузак болуп саналат. Өт кислоталарынын синтези холестерол боор өт кислоталары динин кабыл алат. Билэ кислоталар cholanic кислотасы туунду катары каралышы мүмкүн. сыяктуу Cholanic кислотасы денесинде пайда болгон эмес. холестерол тартып hepatocytes-жылы өт кислоталары негизги түзүлөт - chenodeoxycholic жана cholic. ичеги өсүмдүктөр энзимдердин иш-аракет аркылуу ичегилерден салып өт бөлүү кийин орто Өт кислотасы пайда болот - lithocholic жана deoxycholic. Алар ичегиден-оту менен берилгендиктен, боорго порталы тамырынан канын жана анын абалы менен анда. Өт негизинен өт кислоталары коштолду камтыйт, башкача айтканда. E. кошундулары Glycine же taurine менен анын. 1% жөнүндө өт жана өт кислоталарынын топтолушу. өт бөлүгү катышкан кислоталары gepatoenteralnoy жүгүртүү.Өт кислоталары бир аз бөлүгү - күнүнө 0,5 гр жөнүндө - заъда бөлүнүп. Бул кислота бассейн 10 күндүн ичинде турат өт, боор өт кислотасы менен жаңы синтези менен жабылат. Cholesterol ичеги аркылуу, ошондой эле негизги пайда болот. Ичеги-жылы тамак-аш жана өт курамына боор тартып келет. канга сорулат Cholesterol өт алынган бир бөлүгүн түзөт (ички холестеролбоордо, метионин), ошондой эле тамак-аш алынган бөлчөк (экзогендик холестерол). кыртыштардан холестеролду өчүрдү заъда кийинки чыгарууда менен кошо боор менен өт кислоталары анын кычкылдануу жолу менен пайда болгон (күнүнө 0,5 г жөнүндө) жана өзгөрүүсүз холестерол (ошондой эле миробдорду) чыгарууда менен. туруктуу мамлекеттин ичинде: (CholesterolEnd + Cholesterolзи) - (Cholesterolekskr + Билэ кислоталарekskr) = 0 Бул тең салмактуулук бузулган болсо, кыртыштардын жана кан өзгөрүүлөр холестеролду топтолушу. кан холестерол көлөмүн жогорулатуу - hypercholesterolemia. Бул атеросклероздун Мандела ыктымалдуулугун арттырат. Майлардын метаболизми жөнгө салуу Майлардын метаболизми борбордук толкунданып системасы тарабынан жөнгө салынат. узун терс сезимдерге стресс, Кандагы catecholamines жогорулатуу релизи олуттуу салмак жоготууга себеп болушу мүмкүн. таасири глюкагон catecholamines аракетине окшош lipolytic системасы боюнча. адреналин жана неадреналин плазма май кислоталарын көбөйөт, натыйжада кемик ткань ылдамдыгы липаз жана кыртыш lipolysis ишин жогорулатуу. инсулин Бул lipolysis жана май кислоталарынын тартуу боюнча адреналин жана глюкагон карама-каршы таасир этет. Sth atsenilattsiklazy синтезделишинде азгырат lipolysis стимулдайт. Бул кичинекей бир Hypofunction органы майын депонирлөөгө алып бези (семирүү, кичинекей). Тироксин, жыныстык гормондорОшондой эле көрбөйбүз зат таасирин тийгизет. жаныбарлар менен катар кетирүүчү майын ашыкча алуу себеп болот. Майлардын метаболизми бузулуулар Cholesterol - тирүү организмдердин гана мүнөздүү Стероиддик. Адам денесинде аны түзүүнүн негизги жери - боор, холестерол 50% синтезделет, ичке ичегиде 15-20% түзүлөт, ал калган тери, бөйрөк кабында жана бездерге синтезделет. фонд холестерол жана анын чыгымдардын жолдору булагы Figure 22.1 көрсөтүлгөн. Сүрөт. 22.1. Түзүү жана денедеги холестерол түзүү үчүн бөлүштүрүү. Cholesterol адамдын организми (140 г жөнүндө жалпы суммасы) үч бассейнде бөлүүгө болот: 30 г), bystroobmenivayuschiysya ичеги дубал холестерол, кан плазмасында, боорго же башка parenchymatous органдардын турат, өзгөртүү 30 күн өткөндөн кийин пайда болот (1 г / күн) 50 г), холестерол medlennoobmenivayuschiysya башка органдары жана башка кыртыштардын 60 г), өтө medlennoobmenivayuschiysya MS мээ жана жүлүн, тутумдаштыргыч ткань, өзгөртүү курсу жыл эсептелет. Cholesterol синтези клеткалардын Органеллдер болот. Бул адамдын денеси узун жолдорунун бири болуп саналат. Ал үч этап менен ишке ашат: mevalonic кислота түзүү менен биринчи учтарын, экинчиси - түрү squalene (түзүлүшү түз сызыктуу бир суутек, 30 көмүртек атомдорунан турат). Үчүнчү кадам squalene lanosterol молекуласынын айландырылат учурунда мындан ары холестерол салып сүйлөгөнгө lanosterol 20 жолу катары менен кабыл болот. Айрым кыртыштарда, холестериндин түрү Esters үчүн Гидрокси топ esterified жатат. (: Холестерол acyltransferase atsilKoA) кабыл клеткадагы энзим АСАТ тарабынан кылдат ылгоодон жатат. Колдери жооп да кан липопротеинди пайда, бул жерде бир энзим LCAT (lecithin: холестерол acyltransferase). Cholesterol ТББнын - бул кан клеткалары жеткирүү жана сактоого болгон түрү. кан холестерин 75% жөнүндө Esters түрүндө болот. 3-Гидрокси 3-methylglutaryl-КоА (ГМГ-КоА) - холестерин синтези жөнгө жараянынын негизги энзимдердин ишин жана суммасын таасир тийгизүү аркылуу жүзөгө ашырылат. Бул эки жол менен ишке ашат: 1. phosphorylation / ГМГ-КоА-жылдын dephosphorylation. Инсулин активдүү абалга муну менен аны өзгөртүүгө, ГМГ-КоА-жылдын dephosphorylation өбөлгө түзөт. Ошондуктан, холестерол absorptive мезгил көбөйүп синтези. Бул мезгил ичинде өсүш жана синтезине булагы субстрат бар учурунда - acetyl-КоА. жигерсиз мамлекетке айландырып, белок Креатинкиназа А phosphorylation ГМГ-КоА-дем менен: глюкагон тескери натыйжа берет. Постту absorptive мезгилде холестерол айкалышынын натыйжасы катары менен ачарчылык учурунда бөгөт коёт. 2. ГМГ-КоА-жылдын Inhibition редуктаздык.MS (метаболизм жолдору акыркы продукт), ошону менен өзүнүн синтезин жокко ген ГМГ КоА буйруйт ылдамдыгын төмөндөйт, ушундай эле таасир өндүрүлгөн жана кислоталар өт жатат. кандагы холестерол Транспорт РППУнун бөлүгү катары жүзөгө ашырылат. PL экзогендик холестерол ткандардын эске алууну камсыз кылуу, органдардын жана денеден чыгарууда ортосунда агымын аныктоо. Экзогендик холестерол калган Х курамына боорго жеткирилет. Бар, бирге синтезделген ички холестерин менен жалпы фонд түзөт. hepatocytes ТАГ жана VLDL холестерол-жылы пакеттелген жана ушул түрүндө канга бөлүп жатат. LP липаз астында кан VLDL биринчи LPPP prevraschayutya менен glycerol жана май кислоталарын тегин күйгөндө, анан холестерол жана анын Esters 55% чейин камтыган LDL үчүн. LDL - ал кыртышка салып турган транспорттук холестерол негизги түрү (холестерол жана кан анын Esters 70% LDL бир бөлүгү болуп саналат). боорго кан LDL (75%), ошондой эле алардын жер үстүндөгү LDL кабылдагыч боюнча да башка кыртыштардын. клетканын ичине кирген холестериндин өлчөмү анын зарылдыгын ашса, LDL сезгич синтези кан холестерол агымын төмөндөтөт, бөгөт болууда. клетканын эркин холестерол топтолушун азайтуу менен, тескерисинче, ишке кабылдагыч синтези. LDL кабылдагыч тартылган гормондор айкалышынын жөнгө салуу: инсулин, triiodothyronine жана жыныстык гормондор кабылдагычы пайда көбөйтүүгө жана глюкокортикоиддер - кыскартуу. Ошондуктан, "кайтарым холестерол транспорт" деп аталат, б.а. жолдор, боорго холестерол кайтарылышын камсыз кылуу липопротеинди негизги ролду ойной. Алар боордо олуттуу холестерол жана ТАГ акысыз жетиле прекурсорлорду катары синтезделет. башка PL жана клетка кабыкчасындагы аны алып кан прабабушки HDL холестеролду каныккан. Липопротеинди холестерол өткөрүп берүү бетинде жайгашкан тартылган LCAT энзим болуп саналат. Бул энзим магистр үчүн phosphatidylcholine (lecithin) бир май кислотасы калганды маани берет. Натыйжада бир hydrophobic молекула, ичибизди липопротеинди түрткү болгон холестерол Эстер, болуп саналат. HDL холестерол байытылган Ошентип nezrezye, липопротеинди 3 айланат - жетилген жана ири көлөмү бөлүкчөлөр. HDL 3 TAG холестерин Esters алмашуу VLDL жана LPPP белгилүү бир протеин менен камтылган, липопротеиддердин ортосундагы холестерол Эстер берүүнү. Ошентип HDL 3 LPVP2 өз кезегинде, алардын көлөмү ТАГ топтолушунан улам жогорулайт. LP караштуу VLDL жана LPPP жерге жана боорго холестеролду жеткирүү LDL, динин липаз. холестерин аз бөлүгү боор жана LPVP2 LPPP тапшырылат. Өт кислоталарынын синтези. боор, холестерин күнүнө Өт кислотасы 500-700 мг чейин синтезделет-жылы. Алардын пайда болушу холестерол гидроксилдик топтору каптал чынжыр hydroxylases жана жарым-жартылай кычкылдануу кабыл катышуусу менен кабыл киргизүү камтыйт (Figure 22.2.): Сүрөт. 22.2. Өт кислоталар схемасы. Биринчи синтез жооп - 7-а-gidroksiholesterola калыптандыруу - бул ченемдик болуп саналат. Бул жооп катализаторлору энзимдердин иш-жолу акыркы менен тосулуп турат - zholchnymi кислоталар. Дагы бир механизм энзим (7-а-гидроксилаз активдүү phosphorylated түрүндө) phosphorylation / dephosphorylation жөнгө салуу болуп эсептелет. энзим көлөмүн өзгөртүү менен мүмкүн жана жөнгө салуу: холестерол тайымдарга ген копиялоо 7-а-гидроксилаз жана кислоталар жемелеп, өт. Калкан безинин гормондору 7-а-гидроксилаз синтезделишинде азгыра жана эстроген - жемелеп. Өт кислоталардын синтезине эстроген бул таасир аялдардын өттөгү 3-4 эсе көп эркектерге караганда эмне үчүн түшүндүрөт. деп аталган холестерол жана chenodeoxycholic кислота cholic пайда "баштапкы zholchnymi кислоталары." кирүү молекулалары карбоксил тобу zholchnoy кислотасы Glycine же taurine үчүн - бул аминокислоталар жапырт бурдук дуушар болот. Conjugation Өт кислотасы активдүү түрлөрүн түзүү менен башталат - КоА туунду жана андан кийин taurine же Glycine кошулуп, натыйжада 4 Variant туундулары болуп саналат: taurocholic жана taurohenodezoksiholevaya, glycocholic жана glycochenodeoxycholic кислотасы. Алар баштап Өт кислотасы алда канча күчтүү emulsifiers болуп саналат. денедеги taurine суммасы чектелет бери Glycine менен туундулары, 3 эсе taurine караганда көбүрөөк киреше алып келген. Ичеги-жылы негизги Өт кислотасы туундулары бир аз сандагы enzymatically орто өт кислоталарга бактерияларды өзгөрөт. Deoxycholic кислотасы cholic, lithocholic түзүлгөн жана deoxycholic, аз эрийт жана акырындык менен ичегиде берилип турат. кармап ичегиден өт кислоталарынын 95% жөнүндө, порталына тамырынан аркылуу боорго кайтып келип, андан кийин дагы бир жолу өт кирип бөлүп, жана тоъ майлар, ыштоо-жылы кайрадан колдонулат. Бул жолу кислота enterohepatic жүгүртүүгө өт деп аталат. Заъдан-жылдан бери негизинен орто Өт кислоталары алынып салынат. Cholelithiasis (GSD) - патологиялык жараян магистр негизинде бир пайда т таштар, болгон. өт холестеролду обочолонуп micellar абалда hydrophobic молекулаларды кармап өт кислоталар жана куралы, холестерол катыштык берүү менен коштолууга тийиш. өт боюнча өт кислоталар жана холестериндин катышынын өзгөрүшүнө алып келүүчү себептери: холестерин менен бай азыктар, жогорку калориялуу тамак-аш, т-жылы өт Жаран, кислота синтези оорулар өт, өт суюктугун жугузуунун enterohepatic жүгүртүү бузуу. cholelithiasis менен илдетке чалдыккандардын көбү ал басаъдаган тартып, холестерол ал- суммасында алып жана кислоталар өт салып чыгарат өт өт кислоталардын холестерол жана синтезин синтезин жогорулаган. Натыйжада, MS бара-бара бекемдеп жабышчаак бир тебелеп түзүү, Өттүн боюнча бөлүнүп баштайт. Кээде, белоктор жана кальций туздары билирубиндин импрегнирдик жатат. Stones гана холестерин (холестерин таштар) же холестерин, билирубиндин, протеин жана калий аралашмасы турушу мүмкүн. Cholesterol таштар, адатта, ак жана аралаш болуп - ар түрдүү конус күрөң. түзүү тектердин баштапкы этабында дары chenodeoxycholic кислотасы катары пайдаланылышы мүмкүн. т-жылы бир жолу, ал акырындык менен холестерин таш, бирок бир нече айдан созулган, жай жараян таркатат. Атеросклероз - патология тамыр дубалдын ички бетинде atherogenic кооз табактарда пайда болушу менен мүнөздөлөт. Бул оорунун негизги себептеринин бири - бул денени калтырып, тамак-аш, анын синтези жана чыгарууда тартып холестерол алуунун ортосундагы дисбаланс. атеросклероздун жапа чеккен бейтаптарды, A, D, жана VLDL болгон бирикмелер. Липопротеинди топтоо жана иштеп чыгуу, атеросклероз менен кокустан ортосундагы кайтарым байланыш бар. Бул кыртышында сызыктарын LDL холестерол иштеши түшүнүктөр менен туура келет, липопротеинди - кыртыштардан. Негизги зат "шарт", атеросклероздун өнүктүрүү hypercholesterolemia болуп саналат. (Канда көп холестеролго). hypercholesterolemia иштеп чыгуу: 1. ашыкча холестерол, карбонгидрат, майын улам кирген, 2. Генетикалык шыктуулук, apoB-100 LDL кабылдагычтар же суу чарба курулуштарын турган генетикалык кемчиликтер, ошондой эле apoB-100 өсүшүнө синтезин же затты (үйбүлөлүк айкалышкан hyperlipidaemia учурда, кайда кан топтолуу жана өсүп холестерол жана TAG). атеросклерозу механизмдерин иштеп чыгууда маанилүү ролду LP өзгөртүү ойнойт. LDL менен көрбөйбүз нормалдуу түзүлүшү жана белок курамын өзгөртүү органына аларды чет өлкөлүк кылат, ошондуктан клеткалардын менен алсыздандырып үчүн жеткиликтүү. LP өзгөртүү бир нече механизмдер менен пайда болот: 1. протеиндердин glycosylation, кан глюкоза топтолуу жогорулашы менен кездешет 2. кычкылы өзгөртүү apoB-100 липиддер, липопротеиддердин түзүмүндөгү өзгөрүүлөр алып келет, 3. аутоиммундук комплекстери түзүү PL антитело (PL autoantibodies түзүүнүн себеби болуп калышы мүмкүн барак). согушу менен өзгөртүлүшү LDL арна. Бул жараян клеткаларга анын келген учурда эле алек холестерол суммасында атайын кабылдагычтар аркылуу Бирок subendothelial космоско кайсы кире "көбүк клеткалар", кирип холестерин ашыкча жана кайра согушу тарабынан көзөмөлдөнүп турат. Бул кан тамырлардын дубалына көрбөйбүз жарнактары же сызыктар пайда алып келет. Бул этапта, тамыр эндотелий түзүлүшүн сактай алабыз. эндотелий зыян пайда көбүк клеткалардын санын көбөйтүү менен. Таттыбүбү кошулуу үчүн зыян берет. Натыйжада, Таттыбүбү жыйындысын өбөлгө өздөрү чыгарган thromboxane, ошондой эле жылма булчуң клеткаларынын жайылтпоо стимулдайт, Таттыбүбү-алынган өсүшү таасирин өндүрүп башташат. артериялык дубалдын ички катмары макулуктардын акыркы которгондо, бул тузулуш өсүшүнө көмөк көрсөтүү. Андан ары ошол жерде өсүп тактасында булалуу тканы, арасы мейкиндиктеги сактоого булалуу баштыкча ээгинин жана холестериндин астында клеткалар. тузулуш акыркы этабында кальций туздары менен импрегнирдик жана абдан жыш болуп калды. Тактайда жерлер тиешелүү бөлүгүндө көп идиш Карандаш үлгүлөрүн Тромболитикалык, кан айлануу кескин бузулушу алып түзүлөт жана миокарддын кыртыштарга. холестерол синтези жөнгө салуу - анын негизги энзим (ГМГ КоА) ар кандай жолдор менен созулган. Phosphorylation / dephosphorylation ГМГ-редуктаздык. инсулин үлүшүн бир топ көбөйтүү менен / Бул энзим phosphorylated жана активдүү болуп жатат глюкагон. 2 энзим аркылуу инсулин иш-аракет. Креатинкиназа ГМГ-КоА-phosphatase, алардын privraschaet Креатинкиназа жигердүү dephosphorylated мамлекет: Phosphatase ГМГ-КоА-dephosphorylated жигердүү мамлекет аны айландыруу менен редуктаздык. Эгер ушундай болуп жатса натыйжасы активдүү түрү defosorilirovannoy ГМГ-КоА-түзүү болуп саналат. Ошондуктан, холестерол absorptive мөөнөтү өскөн. Бул мезгил ичинде өсүш жана холестериндин синтезине булагы субстрат бар учурунда - acetyl - КоА (acetyl-КоА-глюкоза жана май кислоталарынын чиришинен пайда болгон эле нан бар карбонгидрат жана майлардын, натыйжада). аркылуу глюкагон пост absorptive абалда proteingenazu А стимулдайт phosphorylation боюнча ГМГ - КоА - редуктаздык, жигердүү эмес мамлекет аны айландыруу. Бул иш-аракеттер да глюкагон бул phosphorylated жигердүү абалда ГМГ-КоА-сактоо, phosphorylation жана phosphatase ГМГ-КоА-жылдын inactivation сигнал улам жардам берет. Постту absorptive орозо мезгилинде холестерол айкалышынын натыйжасы катары жана ички синтези басат. тамак-холестерол мазмуну 2% га чейин эске алынган болсо, анда чукул ички холестерол синтезделишинде азайган. Бирок, холестерол синтездөөгө эч кандай толук токтотуу жок. ар кандай адамдардын тамак холестерол аракетине холестериндин биосинтези тыюу салуу даражасы ар түрдүү болуп саналат. Бул холестерол түзүү жараяндардын инсандардын көрсөтөт. холестерол синтези абалын көрyп азайтуу, ал канга анын топтоо кыскартуу мүмкүн. Эгерде бузулган холестерол тамак- аштын ортосундагы тең салмактуулук жана анын метаболиттери денеде бир тараптан жана өт кислоталары чыгарууда жана холестериндин - башка, Кан менен кыртыштарда холестерол топтолуу өзгөрөт. абдан оор кесепеттерге алып келсе, холестерол жана кан топтолуу жогорулашы менен байланышкан (hypercholesterolemia) бул атеросклероздун жана cholelithiasis ыктымалдыгын жогорулатат. Үй hypercholesterolemia (НСН) - бул түрү көбүрөөк кездешет - 200 кишиге 1 иши жөнүндө. hypercholesterolemia менен тукум куума кемтиги клеткалар тарабынан LDL өздөштүрүү бузуу менен, демек, LDL жана катаболизм кыскартуу болуп саналат. Натыйжада, кан LDL бир LDL кандагы холестеролду, анткени анын өкчөмө таш боюнча топтолушун жогорулатат-жылы. Ошондуктан, артериялардын капталдарында жылы GCN кыртыштарда холестерол мүнөздүү катмарланышы, айрыкча, тери менен (xanthomas) болот. ГМГ-КоА-ажыратуу холестерол зат алмашуу жолун акыркы продукт. ген ГМГ тако ыкма өз синтезин жокко КоА-редуктазы буйруйт ылдамдыгын азайтат. боор жигердүү холестерол, өт кислоталар жана ошондуктан үгүттөө ген ГМГ-КоА ишинен өт кислоталарынын синтези жүрөт. ГМГ-КоА-3 жөнүндө синтези кийин бар, бул энзим холестерол синтези ажыратуу натыйжалуу жөнгө салуу болуп саналат.
|